

初期サッカード視覚運動系のスパイキング
ニューロンレベルでのモデル化

京都大学 ジャン・モーレン(写真)
奈良先端科学技術大学院大学 柴田 智広
沖縄科学技術大学院大学 銅谷 賢治
(脳神経系WG)
巨視的な視覚運動の挙動は、脳内におけるシステム規模の機構間の高いレベルの相互作用に依存しますが、そのような機構の機能的メカニズムは、低いレベルの神経生理に基づくものです。生物の挙動を十分に理解するために、これらの機構について両方の規模で見る必要があります。記述レベルが多角的になることで、構造的な複雑さや結果として生じるモデルの計算要求が急速に増しています。
我々の最終目的は、我々および他のグループと共同で、網膜から眼筋運動系までの完全な知覚行動ループを構築することであり、現在の目的は、特に上丘(SC)に重点を置いた、衝動性眼球運動(サッカード)を生成するニューロンレベルのモデル化です。サッカードは再現性および常同性が高いという点で良い標的となります。
我々は、現在の神経生理学の最高の知識に基づいた、上丘中間層(SGI)における大規模なスパイクニューロンレベルのモデルを構築するためにNESTシミュレーションツールを使用します(図1)。また、NMDAのシナプス入力で増強した、コンダクタンスに基づく順応性の指数関数的なIaFニューロンモデルを使用します。このモデルのニューロンの数はおよそ100kで、入力発生および活動統合の支援ユニットのほか、バースト、ビルドアップ、半視野、強抑制、および抑制介在ニューロンを伴います。SGIは網膜位相で、眼球運動系の最初の部分を形成しています。網膜は、皮質の視覚系だけでなく上丘浅層にも投射します。上丘浅層は局所的に突出した領域を選択し、中間層部の対応する点を直接あるいは間接的経路を介して活性化します。浅層部は知覚領域ですが、中間層部は運動系の部分であり、これらがこの知覚行動ループのセンサーとモーターの集合部を構成しています。
サッカードは、SGIで、突出点の周りに拡散しているニューロンの活性化を増強して惹起されます。ここでは、外側のSGIを単一の抑制入力に単純化していますが、通常は、脳幹や他の領域のニューロンによって抑制されています。外部抑制を放出すると、バーストニューロンが脳橋と中脳内でバーストニューロン系を活性化するスパイクのNMDAシナプス介在性バーストを生成します。バーストニューロンは次々と眼球へ水平および垂直に移動し、小脳によって制御されます。
SGIはかなり解明されていますが、この領域での活性に関しては未だに多くの問題が残っています。バーストは網膜部位の指標や移動ベクター、速度コマンドになり得るものの、ビルドアップニューロン間での活性化拡散の重要性についても明らかにはなっていません。我々は、この領域における既知の神経生理を踏まえつつ、スパイキングニューロンの活性化とビルドアップニューロン間の活性化拡散の最近の研究結果について説明を試ています(図2)。
バーストニューロン活性は、サッカードの距離依存的な眼球運動の特性を経時的に示します。バースト特性は、NMDAシナプス生成バーストと中脳網様体で蓄積したスパイクからの直線的抑制フィードバックにより構築されます。その活性により、バースト特性を実験的に測定することができます。ビルドアップニューロン活性の拡散は、局所的な結合内および抑制的なニューロン間で生じます。我々は、活性の位置ではなく量によって、サッカードの進行を経時的に追跡できることを提唱しています。これにより、上丘深層部でバーストニューロン活性を直接抑制する抑制ニューロンが誘発され、間接的に吻極に制御を回復させます。これがサッカードの終了時期の正確性を高めています。
我々は、上丘浅層の構築や、我々のモデルと、同様に詳細な網膜および脳幹系のモデルとを組み合わせることを目指しています。また、そのモデルを物理的入出力に結合することで具体化し、これによって計算的に重い行動モデルもより自然な設定にできる可能性について探索しているところです。
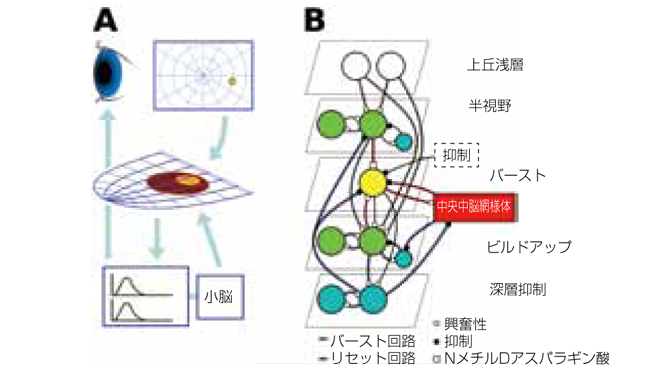
図1:A)皮質下サッカード系の主成分と流れ。網膜からの出力は網膜位相の上丘を活性化し、網膜位相の上丘は脳橋および中脳内の水平方向と垂直方向の眼球運動系を活性化する。小脳は抑制因子として作用する。
B)上丘中間層(SGI)モデルの回路。赤の結線はバースト生成回路、青の結線は活性化の拡散とサッカード系のリセット、グレーの結線は両方に共通の回路を示す。
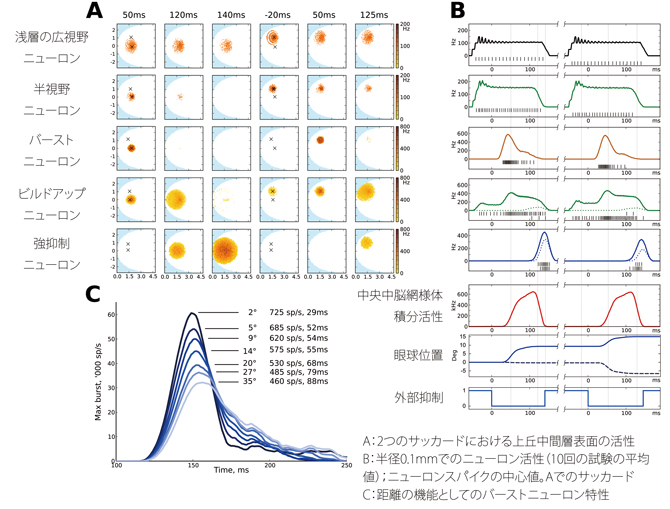
図2:A)半径9°、角度0°の時、半径8°、角度45のサッカードにおけるSGI表面の経時的活性変化。各点は、1モデルのニューロンで平均10msの活性。時間はサッカードの脱抑制に対して相対的。上から、浅層の広視野ニューロン、半視野ニューロン、バーストニューロン、ビルドアップニューロン、強抑制ニューロン。左から、バーストピーク、ビルドアップ活性ピーク、強抑制リセット、第2サッカードの準備活性、第2 バーストピーク、第2ビルドアップピーク。
B)A)での各サッカードの中心点から半径0.1mmでの10回の試験の平均活性。同じ順序で示す。黒のバーは1回の試験でのニューロンスパイクの中心値で、上列は第1サッカード、下列は第2サッカード。各サッカードのビルドアップニューロンと強抑制ニューロンの点線は、もう一方のサッカードでの活性を示す。下に、全cMRF積分活性、水平方向(実線)および垂直方向(破線)の眼球位置の測定値、外部抑制を示す。
C)サッカード距離の機能としてのSGIのバーストニューロン活性の経時的活性変化。サッカード距離が長いほど活性ピークは低下し、持続時間は延びる。
BioSupercomputing Newsletter Vol.6
- SPECIAL INTERVIEW
- 新しい流体構造連成解析手法(ZZ-EFSI)の開発によっていち早く高い演算性能を達成
東京大学大学院工学系研究科 特任准教授 杉山 和靖 - 開発・高度化チームに聞く「京」の実力と高い性能を引き出すために続くチューニングの取り組み
理化学研究所 生命システム研究センター 生命モデリングコア計算分子設計研究グループ
グループディレクター 泰地 真弘人
理化学研究所 次世代計算科学研究開発プログラム 次世代生命体統合シミュレーション研究推進グループ
生命体基盤ソフトウェア開発・高度化チーム 上級研究員 大野 洋介
理化学研究所 HPCI計算生命科学推進プログラム 高度化推進グループ
高度化推進チーム 上級研究員 小山 洋
理化学研究所 次世代計算科学研究開発プログラム 次世代生命体統合シミュレーション研究推進グループ
生命体基盤ソフトウェア開発・高度化チーム 研究員 舛本 現
理化学研究所 次世代計算科学研究開発プログラム 次世代生命体統合シミュレーション研究推進グループ
生命体基盤ソフトウェア開発・高度化チーム リサーチアソシエイト 長谷川 亜樹
- 研究報告
- 全原子分子動力学シミュレーションによる多剤排出トランスポーターAcrBの機能解析
横浜市立大学大学院生命ナノシステム科学研究科 山根 努 / 池口 満徳( 分子スケールWG) - ヒト循環器系のマルチスケールモデリング
理化学研究所 次世代計算科学研究開発プログラム 梁 夫友( 臓器全身スケールWG) - 初期サッカード視覚運動系のスパイキングニューロンレベルでのモデル化
京都大学 ジャン・モーレン
奈良先端科学技術大学院大学 柴田 智広
沖縄科学技術大学院大学 銅谷 賢治
(脳神経系WG) - 大規模並列用MDコアプログラムの開発
理化学研究所 次世代計算科学研究開発プログラム 大野 洋介(開発・高度化T)
- SPECIAL INTERVIEW
- 複雑な生命現象の理解と予測に向けて計算生命科学の明日を拓く
理化学研究所 HPCI計算生命科学推進プログラム プログラムディレクター 柳田 敏雄
理化学研究所 HPCI計算生命科学推進プログラム 副プログラムディレクター 木寺 詔紀
理化学研究所 HPCI計算生命科学推進プログラム 副プログラムディレクター 江口 至洋
- 研究報告
- 全原子モデルにもとづくヌクレオソームポジション変化の自由エネルギープロファイル計算
日本原子力研究開発機構 量子ビーム応用研究部門
河野 秀俊 / 石田 恒 / 米谷 佳晃 / 池部 仁善
(分野1-課題1) - 骨格筋の活動の推定と脊髄反射の神経モデル
東京大学情報理工学系研究科 中村 仁彦(分野1-課題3)
- 報告
- ISLiM 成果報告会2011
理化学研究所 次世代計算科学研究開発プログラム 田村 栄悦 - 高等学校で行った計算生命科学の授業
理化学研究所 HPCI計算生命科学推進プログラム 鎌田 知佐 / 藤原 康広 / 江口 至洋